Рис 2 fed
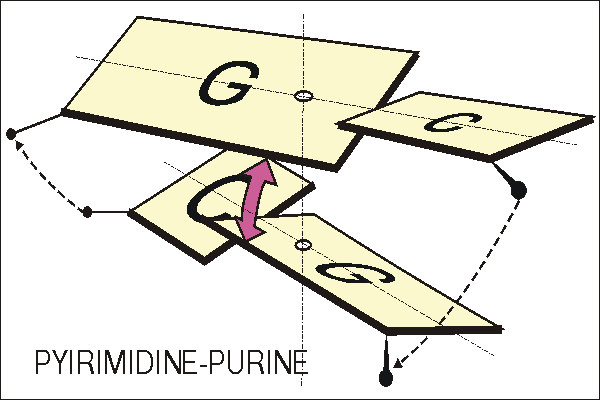
Рис 2-1. Иллюстрация к первой стратегии. Пространственное
столкновение
(по Калладину) пуринов из противоположных нитей, имеющих место в
узкой
бороздке в паре Y-R (Y-пиримидин, R-пурин). Пример приведен для C-G
динуклеотида. В каждой комплементарной C.G паре имеет место
положительный пропеллерный угол относительно большой оси пары. Пурины
из соседних пар оказываются в слишком тесном контакте в малой бороздке
из-за пропеллерного угла. В другой паре R-Y имело бы место
аналогичное
столкновение в широкой бороздке. Калладин предположил, что для
разрешения конфликта пуринов локальные спиральные углы "twist" и
"roll"
адаптируются так, чтобы минимизировать это столкновение пуринов.
Это подтверждается и экспериментально во многих случаях.
(
Calladine, 1982
;
Dickerson, 1983
).
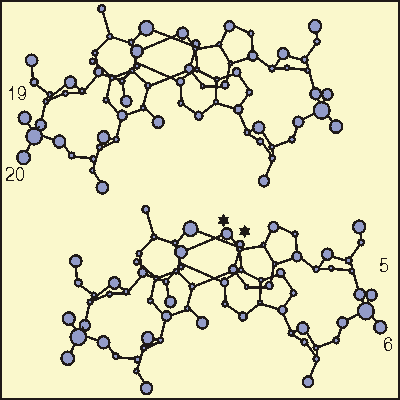
Рис 2-2. Стереорисунок ко второй стратегии разрешения конфликта
пуринов.
для пары оснований A5.T20 и A6.T19 из ДНК додекамера
C-G-C-G-A-A-T-T-C-G-C-G
(Prive et al,1991). Звездочкой указаны аминогруппы при N6
аденина,
которые, согласно модели клиньев, находятся в слишком тесном контакте.
Взаимное отталкивание приводит к раскрытию угла "roll" между
парами
оснований в сторону верхней части рисунка и раскрытию угла "tilt"
в правую сторону. (
Prive et al, 1991
)
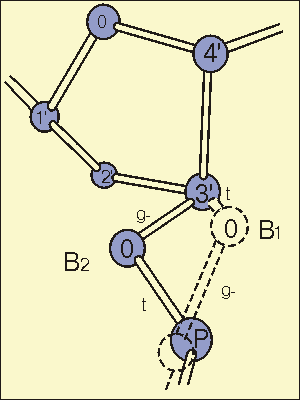
Рис 2-3. Иллюстрация к третьей стратегии разрешения конфликта
пуринов.
Альтернативная конформация сахаро-фосфатной цепи ДНК. В случае
BI
конформации торсионный угол C4'-C3'-O3'-P находится в trans-
конформации, и угол C3'-O3'-P-C5' в gauche- конформации. В случае
BII конформации первая связь в gauche-, а вторая - trans. Вращение
C3'-O3' связи из положения BI в BII разворачивает связь C3'-C2'назад
за плоскость рисунка, предпочитая классическую C2'-endo конформация
B ДНК. Переход из BI в BII приводит к увеличению расстояния
между
парами вдоль оси спирали и изменению других конформационных параметров
для предотвращения конфликта пуринов.
Ссылки на рисунок: