Рис. 14.5(epig). Формирование неоцентромер у человека и мух
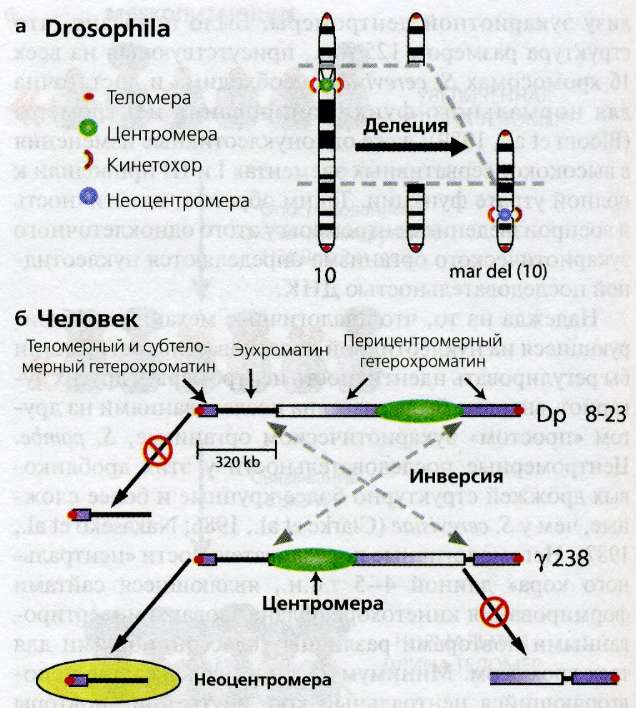
(а) Хромосомы человека, несущие неоцентромеры, которые демонстрируют центромерную функцию и формирование кинтетохора в отсутствие центромерной ДНК, обычно связаны с крупными перестройками ( Amor and Choo, 2002 ). В этом примере происходящая от хромосомы 10 неоцентромера (mar(del)10), структура которой указывает на формирование посредством большой интерстициальной делеции, удалившей эндогенную центромеру (серые пунктирные линии). Mar(del)10 была открыта у индивидуума, в кариотипе которого была также кольцевая хромосома (ring(del)10, не показана), содержавшая ДНК из делетированного участка. Последовательность событий для неоцентромер человека неясна; формирование неоцентромеры могло бы происходить вначале, давая дицентрическую хромосому, которая впоследствии подвергается перестройкам, или же формирование неоцентромеры могло бы происходить после делеции эндогенной центромеры.
(б) У мух неоцентромеры могут создаваться экспериментально из минихромосомы, охарактеризованной на молекулярном уровне. Фрагмент эухроматина величиной 320 т.п.н. и теломерный хроматин, не содержащий центромерной ДНК, можно отделить от остальной минихромосомы, используя облучение. Этот фрагмент, который должен бы быть "ацентрическим", может стать функциональной неоцентромерой, которая надежно воспроизводится в митозе и мейозе и содержит белки центромеры и кинетохора, в норме ограниченные эндогенной центромерой ( Blower and Karpen, 2001 ). Однако для формирования неоцентромеры требуется близость к эндогенной неоцентромере (420 т.п.н.), как в инверсионном деривате гамма238; более того, формирование неоцентромеры не происходит по обе стороны от центромеры, когда присутствует перицентрический гетерохроматин ( Maggert and Karpen, 2001 ). Эти результаты заставляют предполагать, что формирование неоцентромеры происходит через эпигенетическое распространение центромерного хроматина в соседний эухроматин, за которым следует эпигенетическое воспроизведение центромерной идентичности и функции. Блокирование этого процесса гетерохроматином согласуется с наблюдением, что сверхэкспрессированная CENP-A включается эктопически в эухроматин, но не в гетерохроматин ( Heun et al. 2006 ), и позволяет предполагать, что протяженность центромерного хроматина определяется двумя эпигенетическими процессами: загрузкой и распространением CENP-A и формированием-блокированием гетерохроматина.
Ссылки на рисунок: