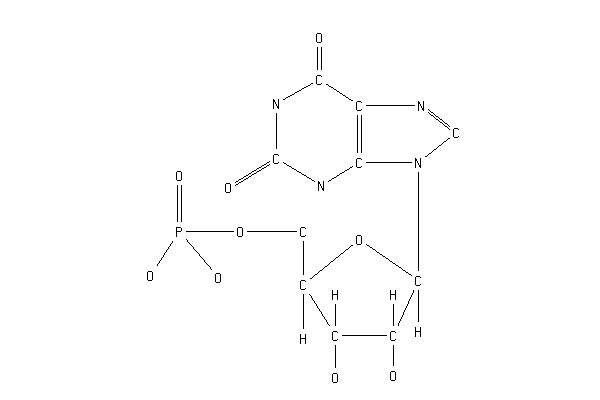
XMP+ATP+Gln=GMP+AMP+Glu+PPi
ATP + XANTOSINE-5-PHOSPHATE + L-GLUTAMINE = АМP +( PPi )+ GМP + L-GLUTAMATE Реакция необратима [ Abrams R.,1959 , Lagerkvist U.,1958 , Franko R.,1984a , Franko R.,1984b ].
Реакция представляет собой последнюю стадию в биосинтезе пуриновых нуклеотидов и катализируется ферментом ГМФ-синтетазой (гидролизирующей глутамин) ( GMPS , ГФМС ).
Тип реакции: перенос амидной группы; аминирование за счет энергии сопряженной реакции расщепления макроэргической связи АТР. [ Schomburg D.,1990 ].
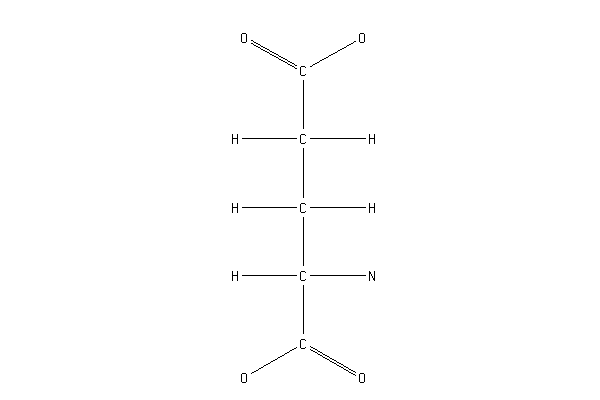
Исследование механизма реакции показало, что реакция упорядоченная, последовательная. 1-м субстратом, который взаимодействует с ферментом, является ХМР, 2-м - АТР, 3-м - NH4+[ Franko R.,1984 ]. По отношению к субстрату ХМР - реакция нулевого порядка [ Abrams R.,1959 ]. Предполагается, что глутамин связывается с SH-группой на ферменте, происходит нуклеофильная атака гамма-карбонильной группы глутамина с освобождением амидогруппы аммония, последний связывается с металлом, присутствующим в активном центре, и переносится к субстрату- акцептору [ Patel N.,1977,a ]. В качестве аминодонора в реакции, катализируемой ГМФС, более предпочтителен L-глутамин, чем NH4+, что характерно для фермента из фибробластов человека [ Page T.,1984 ], из асцитных клеток саркомы Иошида (М) [ Hirai K.,1987 ] и из печени голубя [ Lagerkvist U.,1958 ], но не для ГМФС из E.coli [ Patel N.,1977,b ]. Для ГМФС из печени голубя активность с NH4+ составляла лишь 15% от активности с L-глутамином. Значения Km были равны для ХМР - 0,0046 mM, АТР - 0,12 mM, глутамина (GIn)-0,3 mM, NH4+ - 120 mM для ГМФС из ас- цитных клеток саркомы Иошида [ Hirai K.,1987 ] или 0,0036 mM для ХМР , 0,28 mM для АТР , 68 mM для глутамина ( GIn ) и 36 mM для (NH4)2SO4 для GMPS из асцитных клеток опухоли Эрлиха [ Spector T.,1975 ].
ГМФС обнаружена у человека: в слизистой толстой кишки [ Natsumeda Y.,1985 ), фибробластах , костном мозге , лейкоцитах , эритроцитах , плаценте , печени [ Page T.,1984 ]; млекопитающих: в лимфоцитах (М) [ Marinello E.,1991 ]; в тимусе теленка [ Abrams R., 1959 ]; в мозге крысы [ Achleitner E.,1985 ], почках крысы [ Boritzki T.J.,1981 ; Hinai,1987 ], печени крысы , гепатоме крысы и др. органах крысы [ Boritzki T.J.,1981 ; Hirai K.,1987 ; Weber G.,1985 ; Weber G.,1982 ], асцитных клетках саркомы крысы [ Hirai K.,1987 ; Weber G.,1983 ]; костном мозге кролика [ Meister A.,1973 ]; асцитных клетках карциномы мышей [ Spector T.,1975 ]; у китайского хомячка [ Kaufman E.R.,1985 ]; у птиц: в печени голубя [ Lagerkvist U.,1958 ].
Показано, что реакция катализируется ферментом, локализованным в цитозольной фракции [ Boritzki T.J.,1981 ]. У крыс исследовали распределение активности ГМФС в различных органах [ Hirai K.,1987 ]. Показано, что активность ГМФС убывала в ряду : семенники крысы , мозг, кишечник крысы , тимус крысы (вилочковая железа), яичники крысы , легкие крысы , жировая ткань крысы , сердце крысы , почки, печень, селезенка крысы , скелетные мышцы крысы от 115 до 8,9 мкЕд/мг белка в норме. Наиболее высокая активность была в асцитных клетках саркомы - 175 мкЕд/мг белка. После частичной гепатэктомии и регенерации печени активность ГМФС возрастала и достигала максимума через 24-36ч (180%) после операции, затем постепенно уменьшалась.
п-Хлормеркурийбензоат (РСМВ) ингибировал на 50-100% ГМФС из асцитных клеток Эрлиха печени голубя и из E.coli [ Spector T.,1975 ; Lagerkvist U.,1958 , Patel N.,1977,a ]. Генетические исследования для ГМФС у человека и млекопитающих не известны.
У E.coli гены GMPS и IMP-дегидрогеназы относятся к одному оперону [ Reichert U.,1975 ]. Биосинтез GMP из IMP через ХМР в E.coli К-12 контролируется двумя генами gua A и gua B [ Fukumaki Y.,1977 ].
{kind=link}
{kind=link}
{kind=link}