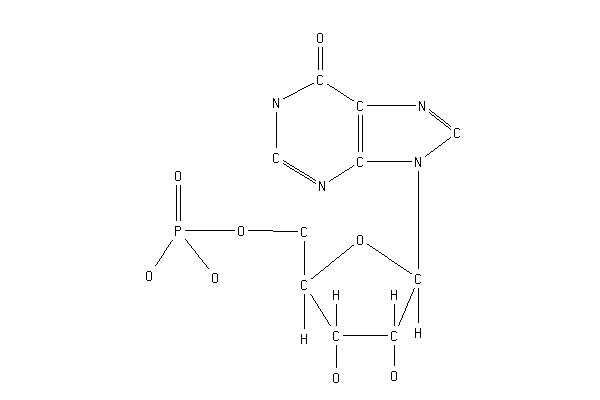
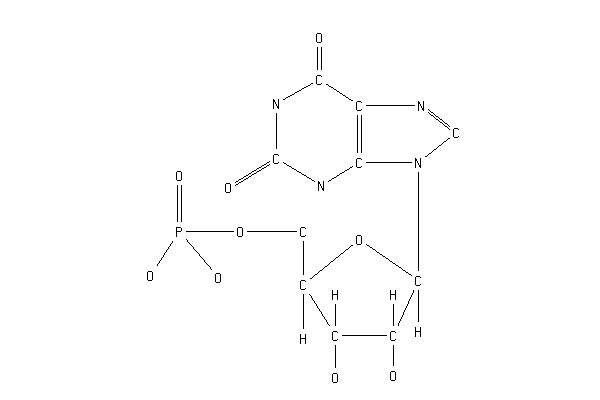
IMP+NAD+H2O=XMP+NADH
INOSINE-5'-PHOSPHATE + NAD +H2O = XANTHOSINE-5'-PHOSPHATE + NADH +H
Реакция не обратима [ Magаsanik B.,1957 ]. Тип-окислительно- восстановительный. Реакцию катализирует IMP-дегидрогеназа .
Фермент обнаружен у бактерий [ Magasanik B.,1957 ] плесневых грибов [ Pourquie J.,1969 ], у птиц [ Nagata K.,1965 ], у млекопитающих [ Tiedeman A A .A.,1991; Shimura K.,1983 ], в том числе и у человека [ Collаrt F.R 1988; Natsumeda Y.,1990 ; Holmes E.W.,1974 ].
У млекопитающих фермент найден в клетках многих органов: печень крысы [ Tiedeman A.A.,1991 ; Hirai K.,1987 ], легкие крысы , сердце крысы , почки крысы , мозг крысы , скелетные мышцы крысы , тонкий кишечник крысы , жировая ткань крысы , семенники крысы , вилочковая железа крысы [ Hirai K.,1987 ], селезенка человека [ Natsumeda Y.,1990 ; Hirai K.,1987 ], лейкоциты [ Nagai M.,1991 ], лимфоциты [ Marinello E.,1991 ] и плацента человека [ Holmes E.W.,1974 ].
Фермент локализован в цитозоле клеток [ Shimura K.,1983 ; Holmes E.W.,1974 ], но мог образовывать седиментационноспособный комплекс с компонентами цитозоля с участием гидрофобных и электро- статических взаимодействий [ Shimura K.,1983 ]. У IMP-дегидрогеназы клеток асцитной саркомы Иошида способность образовывать комплекс с цитозольными компонентами выше, чем для фермента из печени, что свя- зывают с быстрым ростом клеток [ Shimura K.,1983 ].
У млекопитающих, в том числе и человека, обнаружены два изофер- мента IMP-дегидрогеназы: IMP-дегидрогеназа I и IMP-дегидрогеназа II. Изофермент I встречается в нормальных тканях, а изофермент II обнаpужен в опухолевых клетках и его количественное содержание в клетке коррелировало со степенью пролиферации клеток [ Natsumeda Y.,1990 ; Nagai M.,1991 ; Masami M.,1992 ].
Кинетика ферментативной реакции описывается упорядоченным пос- ледовательным механизмом связывания субстратов и освобождения про- дуктов: первым связывается IMP, затем NAD+; NADH первым из продуктов диссоциирует из тройного комплекса,а последним -XMP[ Yamada Y.,1988 ].
Регуляция активности IMP-дегидрогеназы у млекопитающих осущест- вляется изостерически. IMP-дегидрогеназа является ферментом, катали- зирующими лимитирующую стадию пути биосинтеза гуаниновых нуклеотидов [ Weber G.,1983 ; Lui M.S.,1984 ] и активность этого фермента критичес- ки влияет на концентрацию GTP и dGTP. Соотношение потоков IMP на синтез GMP через XMP и на синтез AMP через аденилосукцинат (реакция, катализируемая аденилосукцинатсинтетазой ) зависит от концентраций физиологических ингибиторов и от соотношения и количества изофермен- тов. IMP-дегидрогеназа печени крысы [ Shimura K.,1985 ] и плаценты че- ловека [ Holmes E.W.,1974 ] ингибируется продуктом реакции - XMP, AMP и, в меньшей степени,пиримидиновыми нуклеотидами. Ингибирование XMP, очевидно, не имеет физиологического смысла, поскольку в печени крысы и различных гепатомах его концентрация на порядок ниже, чем величина Ki [ Jackson R.C.,1980 ], вследствие высокой активности GMP-синтетазы (гидролизующей глютамин). Найдено, что активность IMP-дегидрогеназы линии клеток мыши L-127 находится под контролем клеточного белка, антионкогена, [ Sherely J.L.,1991 ].
Содержание фермента у крысы [ Hirai K.,1987 ] было высоким в ви- лочковой железе, селезенке и семенниках (в 5-9 раз выше, чем в пече- ни), средним в сердце, почках, печени и легких и низким в скелетных мышцах ( в 2 раза ниже, чем в печени). Содержание фермента резко повышено ( в 8-15 раз) по сравнению с нормальными тканями во всех исследованных гепатомах печени [ Jackson R.C.,1984; Jackson R.C.,1977 ] в лейкозных клетках человека и у пациентов с миелоидным лейкозом [ Yamada Y.,1990 ]. Следовательно, в раковых клетках протекает преиму- щественное использование (туннелирование) IMP для биосинтеза гуани- новых нуклеотидов [ Natsumeda Y.,1988 ], вызывая дисбаланс адениновых и гуаниновых нуклеотидов [ Lucas D.L.,1983 ]. Увеличение содержания IMP-дегидрогеназы в раковых клетках обусловлено синтезом изофермента II, содержание которого в нормальных тканях было низким и резко по- вышалось в процессе пролиферации клеток и коррелировало со скоростью роста клеток;его содержание уменьшалось в созревших pаковых клетках и резко уменьшалось в процессе диффеpенциации клеток [ Natsumeda Y., 1990 ; Nagai M.,1991 ; Masami N.,1992 ]. Изофермент I является консти- тутивным и его содержание оставалось постоянным в нормальных и рако- вых клетках [ Masami N.,1992 ].Содержание IMP-дегидрогеназы увеличива- лось в регенерирующей печени достигая максимума через 18-24 часа [ Shimura K.,1983 ; Jackson R.C.,1977 ]. Использование изофермента IMP- -дегидрогеназы II, как мишени, при поиске специфических антираковых химотерапевтических препаратов очень актуально.
Предполагается, что у млекопитающих IMP-дегидрогеназа кодируется двумя структурными генами (гены IMP-дегидрогеназы) [ Natsumeda Y., 1990 ].
{kind=link}
{kind=link}