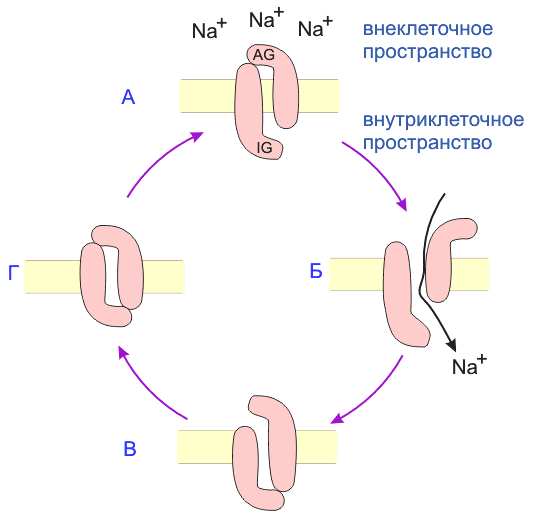
Энхансер: введение
Энхансер (enhancer) [англ. enhancer — усилитель] — цис-действующая регуляторная нуклеотидная последовательность, которая повышает (усиливает) экспрессию генов и может функционировать в разной ориентации и в любых положениях относительно промотора (см. Промотор). Э. локализован обычно в области, расположенной в 5'-положении относительно гена, но может также быть локализован внутри генов (в интронах) и в З'-фланкирующих нуклеотидных последовательностях генов. Для функционирования Э. требуются соответствующие трансдействующие факторы. В отличие от промотора, Э. сам по себе не может обеспечить транскрипцию гена.
Энхансеры представляют собой протяженные последовательности нуклеотидов, которые содержат сайты связывания нескольких факторов транскрипции. Характерными свойствами энхансера являются его способность осуществлять регуляторное действие на промотор на больших расстояниях от него (более 60 т.п.о.), независимость его активности от ориентации по отношению к промоторам и от расположения относительно регулируемого гена.
На рис. I.30 приведены схемы строения нескольких эукариотических генов, отражающие взаимное расположение структурных и регуляторных частей. У гена альбумина ( рис. I.30,а ) энхансер находится перед промотором, а регуляторная часть предшествует его структурной части. У генов иммуноглобулинов регуляторные элементы локализованы в интронах самого гена ( рис. I.30,б ). Энхансер может быть расположен и ниже гена на значительном от него расстоянии, как у бета-глобинового гена ( рис. I.30,в ).
Энхансеры являются своеобразными матрицами для сборки сложных белковых комплексов, структура которых обеспечивает высокоспецифические белок-белковые связывания и передачу регуляторных сигналов РНК-полимеразе II , находящейся в составе инициационного комплекса.
Энхансер ( Grosschedl ea, 1980 ) это генетический цис-элемент, обладающий усиливающим транскрипцию действием, которое практически не зависит от расположения элемента относительно контролируемого им гена. Энхансеры представлены короткими последовательностями ДНК, состоящими из отдельных элементов (модулей), включающих десятки нуклеотидных пар. Модули могут представлять собой повторяющиеся единицы ( Herr ea, 1986 ). Энхансер увеличивает эффективность транскрипции гена в десятки и сотни раз. Особенность энхансеров состоит в том, что они способны действовать на больших расстояниях (более чем 1000 п.н.) и вне зависимости от ориентации по отношению к направлению транскрипции гена. Энхансеры могут располагаться как на 5'-, так и на 3'-конце фрагмента ДНК, включающего ген, а также в составе интронов. Энхансеры обнаружены на 3'-фланге гена, кодирующего полипептидный гормон - плацентарный лактоген человека, а также, в составе интронов генов иммуноглобулинов и коллагена. Энхансеры - основное средство регуляции транскрипции ( Gluzman ea, 1983 ; Khoury ea,1983 ) в клетках высших эукариот.
Энхансеры были первоначально определены, как цис-действующие элементы, которые усиливают транскрипцию независимо от их ориентации и расстояния от гена. Например энхансер "wing margin" cut локуса Дрозофилы находится на расстоянии 85 кб перед промотором, тогда как энхансер мышиного иммуноглобулинового гена Hмю находится в одном из интронов гена. Энхансер альфа цепи Т-клеточного рецептора находится на расстоянии 69 кб в 3'-сторону от промотора. Более того, спаривание хромосом позволяет иногда активному энхансеру на одной хромосоме активировать транскрипцию аллельного промотора на другой[ Blackwood ea 1998 ].
Еще одной важной характеристикой энхансера является его способность активировать любой промотор, расположенный сравнительно недалеко. В целом можно сказать, что энхансер действует на ближайший к нему промотор. Эффект регуляции достигается тем же путем, что и регуляция проксимальными элементами промоторов, т.е. путем сборки на последовательностях энхансера белкового комплекса, который иногда называют энхансеосомой [ Werner ea 1997 ], и его взаимодействия с основным транскрипционным комплексом путем белок-белковых взаимодействий. Ясно, что для того чтобы осуществить эти контакты ДНК, должна изогнуться . Исходно существовало представление, что энхансеры действуют вне зависимости от ориентации относительно промоторов.
В последнее время появились данные, показывающие, что существует стереоспецифичность во взаимодействии энхансерных белковых комплексов, и что благодаря этой стереоспецифичности небезразлично, как направлены многие энхансеры по отношению к промотору, который они регулируют. Неясно, насколько эффект стереоспецифичности распространен, но сам факт ее существования, хотя бы в ряде случаев, говорит о том, что основная концепция независимости эффекта энхансера от ориентации не является универсальной.
Контрольные области генов ( LCR ) часто содержат множественные автономные энхансерные модули которые варьируют в размерах от 50 по до 1,5 кб [ Blackwood ea 1998 ]. Каждый из этих модулей предназначен для выполнения определенных функций. Он может активировать ген на определнной стадии развития или в определенном типе клеток. Отдельный ген может таким образом содержать множество энхансерных модулей, каждый из которых вносит свой вклад в пространсвенную и временную регуляцию экспрессии гена. Вслед за авторами обзора [ Blackwood ea 1998 ] здесь под энхансером я буду подразумевать отдельный модуль. Часто энхансеры содержат те же модули, что и промоторы.
У вируса простого герпеса (ВПГ) обнаружен транскрипционный фактор VP16 , участвующий в активации его непосредственно-ранних генов и присутствующий в зрелом вирионе в количестве около 1000 копий. Белок VP16 взаимодействует с энхансерами, расположенными перед каждым из непосредственно-ранних генов вируса, что приводит к активации их транскрипции. Каждый из энхансеров содержит одну или несколько копий цис-регуляторных последовательностей - нонануклеотидную последовательность 5'-TAATGARAT-3' ( "tat-garat" ) и прямые повторы гексануклеотидной последовательности 5'-CGGAAR-3' ( "cigar" ). В очищенном состоянии белок VP16 не взаимодействует ни с одной из них. В присутствии белковых факторов экстрактов ядер клеток-хозяев VP16 входит в состав многокомпонентного белкового комплекса, формирующегося на последовательности "tat-garat" ( рис. I.31 ). Сходство последовательностей нуклеотидов мотива "tat-garat" и сайтов связывания Oct-белков , обнаруженных в регуляторных последовательностях генов гистонов и иммуноглобулинов , заставило проверить эти белки на способность взаимодействовать с ВПГ-энхансерами. Оказалось, что Oct-1 является вторым белковым компонентом, соединяющимся с "tat- garat"-последовательностью энхансера и облегчающим связывание с ним белка VP16.
Эффективное образование белкового комплекса требует участия других компонентов клетки-хозяина, поскольку очищенные белки Oct- 1, VP16 и последовательность "tat-garat" формируют стабильный комплекс только в присутствии экстрактов ядер клеток HeLa . Белковый фактор, названный HCF , может непосредственно взаимодействовать с VP16 в отсутствие других компонентов комплекса. С помощью мутационного анализа установлено, что 5'- концевой сегмент "tat-garat" ( последовательность "tat" ) необходим для связывания белка Oct-1, тогда как ее 3'-концевая последовательность ( "garat" ) требуется для образования полного белкового комплекса Oct-1-VP16-HCF.
Таким образом, четыре молекулярных поверхности (интерфейса) участвуют в формировании функционально активного комплекса на матричной поверхности мотива "tat-garat" энхансеров непосредственно-ранних генов ВПГ. Одна группа контактов образуется между белками Oct-1 и VP16, вторая - между VP16 и HCF, третья - между Oct-1 и последовательностью "tat" и четвертая - между VP16 и последовательностью "garat" ( рис. I.31 ).
Сложная серия молекулярных интерфейсов участвует в формировании функционального макромолекулярного комплекса на мотиве "cigar" энхансера непосредственно-ранних генов ВПГ. Белок, взаимодействующий с этим мотивом, выделен из экстрактов ядер печени крыс и получил название GABP (GA-binding protein) . Он состоит из двух субъединиц - GABPальфа и GABPбета . Белковые комплексы, похожие на GABP, выделены и из культивируемых клеток человека. Часть аминокислотной последовательности GABPальфа оказалась гомологичной участку ДНК-связывающего домена транскрипционных факторов, принадлежащих к семейству ETS . Все представители этого семейства обладают последовательностью из 85 аминокислотных остатков, которая необходима для их специфического взаимодействия с ДНК. Таким образом, ETS-домен белка GABPальфа образует первый молекулярный интерфейс, обеспечивающий связывание белка с мотивом "cigar" энхансера.
В смеси с GABPбета альфа-субъединица образует белковый комплекс, который обладает бoльшим сродством к мотиву "cigar" энхансера, чем сама альфа-субъединица. Анализ особенностей взаимодействия GABP-комплекса с энхансером позволил обнаружить три высокоспецифичных молекулярных интерфейса: домен полипептида GABPальфа, необходимый для его взаимодействия с белком GABPбета и включающий ETS-домен, и последовательность из 35 С-концевых аминокислотных остатков GAPBальфа. У полипептидной цепи GABPбета во взаимодействии с GABPальфа принимают участие четыре тандемных повтора, состоящие из 33 аминокислотных остатков каждый и расположенные в N-концевой части белка. Анализ аминокислотных последовательностей, гомологичных этому повтору у ряда других белков, позволил выявить высококонсервативную последовательность TPLH . Дополнительные молекулярные взаимодействия между белковым комплексом и последовательностью нуклеотидов "cigar" осуществляются через повторы TPLH после того, как они образуют требуемый комплекс с GABPальфа. При этом аминокислотные последовательности повторов непосредственно контактируют с ДНК.
Наконец, в последнем из известных контактов, имеющих место после сборки белкового комплекса на последовательности нуклеотидов "cigar", участвует так называемый домен димеризации полипептидной цепи GABPбета, расположенный вблизи ее С-конца. Очищенные субъединицы GABPбета в растворе ассоциируют с образованием гомодимеров, которые в присутствии субъединиц GABPальфа формируют гетеродимерный комплекс альфа2бета2, хорошо приспособленный для взаимодействия с гексануклеотидным повтором "cigar" (CGGAAR)2 энхансера.
Анализируя все вышеперечисленные компоненты энхансеров непосредственно-ранних генов ВПГ, можно сделать вывод, что в их активации принимают участие не менее пяти белков (VP16, Oct-1, HCF, GABPальфа и GABPбета), которые участвуют, как минимум, в пяти белок-белковых контактах и шести специфических контактах белок-ДНК ( рис. I.31 ). Прежде всего, обращает на себя внимание кооперативность взаимодействия компонентов, активирующих энхансеры. Ключевые белки этого комплекса - Oct-1, VP-16 и GABPальфа сами по себе слабо взаимодействуют со специфическими последовательностями ДНК, но обладают способностью собираться в стабильный функционально активный комплекс двумя путями: во- первых, объединяясь с кофакторами, которые самостоятельно не способны взаимодействовать с ДНК (например, VP16 с HCF и GABPальфа с GABPбета); во-вторых, контактируя с последовательностью нуклеотидов энхансера, содержащей функциональные участки "tat" и "garat", а также повтор "cigar" и образующей матрицу для геометрически правильной сборки всего активирующего комплекса.
Смотрите также: