Коррекция неспаренных оснований (КНО) в ДНК: введение
Неспаренные основания в ДНК могут возникать в результате трех событий:
1) прямого повреждения оснований ДНК или их предшественников продуктами клеточного метаболизма;
2) ошибочной подстановки некомплементарного основания ДНК-полимеразой в ходе репликации
3) рекомбинационной интеграции однонитевого участка ДНК в неабсолютно идентичную ДНК, партнера по рекомбинации.
Все события приведут к образованию гетеродуплексной ДНК , которая и становится субстратом для ферментов, корректирующих неправильные пары оснований.
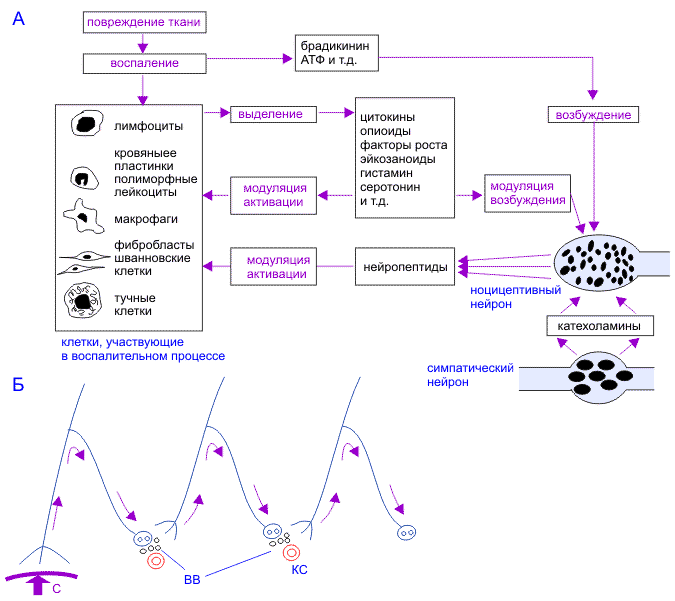
Процесс репарации ДНК при обнаружении ферментами неканонической пары в дуплексе ДНК проходит через ряд реакций.
Обычно такая цепь реакций начинается с обнаружения и удаления одного из нуклеиновых оснований неканонической пары в ДНК, катализируемого соответствующим ферментом группы AP-эндонуклеаз типа II (апуриновых/апиримидиновых эндонуклеаз) [ Kornberg ea 1982 , Краевский ea 1986 ].
Затем происходит гидролитическое расщепление 3'-фосфоэфирной связи, причем фосфат остается в 5'-положении уходящей цепи ДНК. Далее с 3'-концом в месте расщепления цепи связывается ДНК-полимераза бета , приводящая к образованию шиффа между ферментом и 3'-цепью ДНК (реакция А, Рис. 8 ), катализирующая гидролитическое удаление дезоксирибозилфосфатного остатка (реакция В), таким образом освобождая 3'-конец для включения соответствующего канонической паре нуклеотида.
Химический механизм реакции гидролитического удаления включает образование основания Шиффа между остатком Lys72 и альдегидной группой 3'-гликозидного остатка. Далее, после включения в освободившееся положение соответствующего нуклеотидного остатка, ДНК-лигаза воссоединяет непрерывную цепь ДНК.
Наряду с описанным вариантом репарации известны и репараионные процессы, восстанавливающие повреждения одной из цепей ДНК, возникающие в результате мутагенного, лучевого и других воздействий. При этом, по-видимому, во всех случаях заключительные стадии заполнения малых брешей катализируются ДНК-полимеразой бета.
Рентгеноструктурный анализ комплексов [дДНК + ДНК-полимераза] и [дДНК + ДНК-полимераза+dNTP] показывает, что конформация ДНК-полимеразы бета в них сильно различается [ Beard ea 1998 ]. При этом стадией, лимитирующей скорость процесса элонгации является конформационное изменение, превращающее непродуктивный комплекс в продуктивный. Более того, такое превращение эффективно проходит лишь при образовании канонической комплементарной пары, вызывающей конформационное изменение за счет аллостерического влияния. Именно этот механизм главным образом обеспечивает точность синтеза, катализируемого ДНК-полимеразой бета.
В качестве другого примера ДНК-полимераз следует назвать обратные транскриптазы ретровирусов и, вероятно, вируса гепатита В.
Принято различать 2 системы КНО : с длинным ресинтезируемым трактом ( ДКНО , LPMR , long patch mismatch repair ) и
очень коротким трактом ( ОККНО , VSPMR , very short patch mismatch repair ).
Смотрите также: